
The requested URL /topic.htm was not found on this server.
Различают функцию полукружных каналов и функцию отолитового аппарата.
Flourens (1824 г.) первый ввел в изучение физиологии полукружных каналов метод изолированного выключения исследуемого органа.
Перерезая горизонтальные полукружные каналы голубя, он наблюдал полную потерю равновесия и колебательные движения головы в горизонтальной плоскости. При попытке к движению животное падало, вращаясь вокруг продольной оси. При перерезке сагиттальных каналов у голубя появлялись качательные движения головы в сагиттальной плоскости (вверх, вниз), а при попытке к движению животное опрокидывалось на спину, описывая круг в вертикальной плоскости. Перерезывание фронтальных каналов вызывало подобное же явление, но с наклонностью падать вперед. На основании своих опытов Flourens пришел к следующему выводу: со стороны полукружных каналов, расположенных в трех взаимно - перпендикулярных плоскостях, происходит координация движений, сохранение равновесия, ориентация в пространстве. Результаты экспериментов Flourens'a были проверены и подтверждены его современниками (Harless, Chevreul, Czermak, Vulpian).
Если исследованиями Flourens'a было доказано, что полукружные каналы являются органом равновесия, то вопрос о том, что же является нормальным раздражителем полукружных каналов и как нужно представлять работу каналов при нормальных условиях, оставался неразрешенным. Goltz первый дает ответ на этот вопрос. Приписывая полукружным каналам роль периферического органа равновесия, он считает, что нормальным раздражителем вестибулярного нерва является эндолимфа, которая производит давление и растяжение стенок ампулл, сила же давления и растяжения может изменяться в зависимости от изменения положения головы, т. к. высота столба жидкости над ампуллами тогда будет неодинаковой. Получаемыми при этом бессознательными ощущениями и определяется, прежде всего, положение головы, а затем уже при ее посредстве регулируются все остальные движения, необходимые для поддержания равновесия тела.
Это предположение Goltz'a оставалось гипотетическим до тех пор, пока оно не было подтверждено экспериментами Mach'a, Breuer'a и Ewald'a.
Mach в своих опытах с вращением на центрофуге животных и человека пришел к тому выводу, что раздражение ампуллярных нервов при вращении обусловливается изменяющимся давлением эндолимфы, причем последнее зависит не от скорости вращения, а от углового ускорения.
Более убедительными были опыты Breuer'a. Он обнажал перепончатый полукружный канал голубя и выдувал на него из пипетки струю воздуха. Тогда наступало качательное движение головы и глаз всегда в плоскости экспериментируемого канала. Если же, наоборот, воздух всасывался пипеткой, то наступали такие же движения головы и глаз, но направленные в противоположную сторону. Breuer считал, что при такой постановке опыта в полукружном канале образуется ток эндолимфы, в одном случае от простого конца к ампулле, в другом - наоборот.
Еще более убедительными и безупречными в смысле техники считаются опыты Ewald'a. Запломбировав в одном месте наглухо полукружный канал голубя и надавливая особым пневматическим молоточком на участок канала, лежащий между пломбой и ампуллой, он вызывал ток эндолимфы по направлению к ампулле. При этом происходило подергивание глаз в сторону пломбированного канала. При прекращении давления молоточком эндолимфа устремлялась от ампуллы к простому концу, и тогда появлялось движение глаз в противоположном направлении. Несмотря на эти, казалось бы, вполне доказательные, опыты Ewald'a, вопрос о том, что же происходит в полукружных каналах - изменение ли давления или ток эндолимфы - оставался неясным, а между тем решение этого вопроса имело большое значение для клинической лабиринтологии.
Противники Ewald'a и Breuer'a (Biehl, Spitzer, Bard, Wittmaack) говорили, что в капиллярных трубках не может возникнуть токов эндолимфы. Однако, математические вычисления Goede и экспериментальные исследования Mayer'a, Lion'a и Ундрица показали обратное. Последние авторы конструировали капиллярные стеклянные трубки и заполняли их взвесью ликоподия или красных кровяных шариков. При вращении подобных моделей, под микроскопом можно было отчетливо видеть ток взвеси.
Резюмируя экспериментальные данные Flourens'a, Mach'a, Breuer'a, Ewald'a, Mayer'a, Lion'a и Ундрица, можно сказать, что полукружные каналы действительно являются органом равновесия и что специфическим раздражителем окончаний ампуллярных нервов является угловое ускорение, возникающее при поворотах головы или всего тела -и вызывающее токи эндолимфы, которые механически раздражают эти окончания.
Рефлексы полукружных каналов. Из анатомического очерка известно, что полукружные каналы расположены в трех взаимно -перпендикулярных плоскостях. Если человек вращается в одной из этих плоскостей, то произойдет раздражение соответствующих полукружных каналов. Например, при вращении вокруг вертикальной оси (на стуле Вагапу) в плоскость вращения будут попадать преимущественно горизонтальные каналы; при вращении вокруг той же оси, но при наклоне головы вперед, в горизонтальной плоскости вращения будут установлены фронтальные каналы, и угловое ускорение в таком положении вызовет преимущественное раздражение этих каналов; наконец, при вращении около вертикальной оси с наклоненной на плечо головой в плоскость вращения будут попадать преимущественно сагиттальные каналы, и maximum раздражения получат именно эти каналы. Спрашивается, что же произойдет с человеком, если начнет действовать внешний раздражитель полукружных каналов - угловое ускорение? Появится, как было указано, ток эндолимфы, которая отклонит купулу полукружных каналов, и это движение купулы будет вызывать раздражение окончаний ампуллярного нерва. Как только появилось это раздражение, тотчас же появится целый ряд рефлексов в виде изменения тонуса поперечнополосатой и гладкой мускулатуры. Дуги этих рефлексов состоят из афферентных нейронов - n. vestibularis с его ампуллярными окончаниями, коннекторных - вестибулярные центры продолговатого мозга (центры Deiters'a, Бехтерева, Schwalbe, Flechsig'a) и эфферентных- двигательные соматические и вегетативные нервы, снабжающие всю поперечно-полосатую и гладкую мускулатуру.
Рефлексы на поперечно-полосатую мускулатуру (соматические). Среди рефлексов на поперечно - полосатую мускулатуру различают рефлексы: 1) на мышцы глаз, 2) мышцы верхних и нижних конечностей и 3) мышцы туловища. а. Рефлекс на мышцы глаз (нистагм). Если наблюдать движение глаз человека, смотрящего в окно движущегося железнодорожного вагона, то можно заметить, что глаза совершают ряд быстрых ритмических подергиваний, из которых каждое состоит из двух движений - одного быстрого, направленного в сторону движения поезда,и другого-медленного, направленного в противоположную сторону. Такое движение глаз называется зрительным нистагмом и представляет собой весьма целесообразный рефлекс аккомодации глаза к движению. Смысл этого рефлекса заключается в следующем: предположим, что поезд движется вправо, и, следовательно, все предметы, находящиеся вне вагона, будут казаться уходящими влево; далее, представим себе, что человек, смотрящий в окно, фиксировал глазами передвигающиеся влево предметы и провожает их, поворачивая глаза в сторону движения предметов, то-есть влево; наступает момент, когда необходимо будет перенести фиксацию на следующую группу предметов, надвигающуюся справа, т.е. придется повернуть глаза вправо; это движение должно быть быстрым, иначе некоторые предметы успеют проскочить мимо, ускользнуть от фиксации. Первый поворот глаз (влево) называется медленным компонентом, а второй (вправо) - быстрым. Повторяя эти движения, глаза получают возможность рассмотреть наибольшее число мелькающих предметов ("аккомодация глаз к движению").
При наблюдении зрительного нистагма удается установить следующую законность: быстрый компонент нистагма бывает всегда направлен в сторону движения. Совершенно такой же нистагм, подчиняющийся той же законности, бывает и в том случае, если мы будем вращать человека (напр., на стуле Barany) вокруг его вертикальной оси в полной темноте, когда никаких зрительных раздражений не бывает. Такого рода нистагм уже может не зависеть от раздражения сетчатки глаза и, следовательно, он обусловлен исключительно токами эндолимфы в полукружных каналах, т.е. является чисто лабиринтным. Соответственная рефлекторная дуга начинается в окончаниях вестибулярного нерва и далее идет ко дну 4-го желудочка, к area acustica, к вентральному отделу nucleus medialis, а оттуда по tractus vestibulo - longitudinalis к ядрам III, IV, VI пар черепных нервов. Далее импульс распространяется по центробежным путям к мышцам глаза.
По мнению Barany, только медленный компонент зависит от функции упомянутой дуги, быстрый же зависит от деятельности субкортикальных центров. Это доказывается тем, что во время наркоза, когда выключаются субкортикальные центры, быстрый компонент нистагма пропадает и остается один медленный в виде отклонения глаз. Лабиринтный нистагм как бы подкрепляет нистагм зрительный и с этой точки зрения является целесообразным. Происхождение его, по Воячеку, следует объяснить так известно, что при поворотах головы или всего тела происходит кажущееся движение окружающих предметов в сторону, противоположную повороту. При этом одновременно происходит раздражение сетчатки глаза движущимися предметами и раздражение вращением полукружных каналов. Вследствие постоянного одновременного совпадения обоих раздражителей, постепенно, может быть даже в целом ряде поколений, вырабатывалась связь, наподобие условных рефлексов, благодаря которой мало-по-малу раздражение одних лишь полукружных каналов стало давать тот же нистагм, который прежде возникал исключительно при раздражении сетчатки. Эта гипотеза о происхождении вестибулярного нистагма может быть подтверждена следующим опытом. Если наблюдать за нистагмом, вызванным вращением субъекта с закрытыми глазами, когда участвуют одни полукружные каналы, и за нистагмом при таком же опыте, но с открытыми глазами, когда участвуют полукружные каналы и сетчатка глаз, то оказывается, что во втором опыте нистагм бывает менее продолжительным, т.е. зрительный нистагм как бы тормозит вестибулярный. Приведенный опыт позволяет предполагать существование ассоциации между центрами обоих нистагмов. Эта связь касается, главным образом, центров, заведующих быстрым компонентом нистагмов (Хилов).
В зависимости от участия того или иного полукружного канала и нистагм бывает разнообразным. При раздражении горизонтальных каналов нистагм бывает в горизонтальной плоскости, т.е. глаза подергиваются справа налево и слева направо около вертикальной оси. В случае раздражения фронтальных каналов наблюдается ротаторный нистагм, когда глаза совершают вращательное движение вокруг передне-задней оси глаза. При раздражении сагиттальных каналов нистагм бывает вертикальный, т.е. глаза подергиваются снизу вверх и наоборот - около горизонтальной фронтальной оси. Бывает и диагональный нистагм, когда движение глаз происходит в направлении, среднем между вертикальным и горизонтальным. Такой нистагм бывает обычно в случае одновременного раздражения сагиттальных и горизонтальных каналов. Наконец, различают еще смешанные нистагмы, напр.: ротаторно-горизонтальный, ротаторно-вертикальный или вертикально-горизонтальный, которые зависят от одновременного одинакового участия фронтальных и горизонтальных, фронтальных и сагиттальных, сагиттальных и горизонтальных или даже всех каналов.
Встречающиеся варианты нистагма позволяют вывести следующее заключение: нистагм глаза бывает всегда в плоскости, параллельной тем каналам, которые приходят в раздражение. Если участвуют два или все три канала, то плоскость нистагма является результирующей по закону сложения сил. Направление нистагма условно обозначается по направлению его быстрого компонента, т.е. если говорят, что горизонтальный нистагм бьет вправо, то это значит, что быстрые подергивания глаз происходят вправо. При ротаторном нистагме для определения направления судят по быстрому движению верхнего полюса роговицы, а именно, если верхний полюс роговицы совершает быстрое движение вправо, то говорят о ротаторном нистагме вправо.
Нистагм бывает наиболее отчетливо выражен при взгляде исследуемого в сторону быстрого компонента, поэтому при наблюдении за нистагмом просят исследуемого смотреть на палец исследующего, установленный на стороне быстрого компонента. Чтобы узнать, в какую сторону будет направлен быстрый компонент нистагма, руководствуются законом Ewald'a. Ewald экспериментом на горизонтальном полукружном канале голубя установил, что быстрый компонент нистагма всегда направлен в сторону экспериментируемого канала при токе эндолимфы к ампулле. Если же ток эндолимфы идет от ампуллы к простому концу, то нистагм бьет в сторону, противоположную этому каналу. Кроме того, при токе эндолимфы к ампулле нистагм бывает всегда продолжительнее и наиболее отчетлив, чем при токе от ампуллы к простому концу. В отношении фронтального канала - картина обратная. Здесь ток эндолимфы к ампулле вызывает нистагм не в сторону этого канала, как это имело место в опыте с горизонтальным каналом, а наоборот - в противоположную. Так как в дальнейшем при детальном разборе механизма нистагмов придется часто пользоваться законом Ewald'a, то вполне уместно представить его в виде рабочей схемы:
| Правый горизонтальный канал (голубя) | Результат | Левый горизонтальный канал | Результат |
| Ток эндолимфы направлен к ампулле | Нистагм направлен вправо, горизонтальн., продолжительный | Ток эндолимфы направлен к ампулле | Нистагм направлен влево, горизонтальн., продолжительный |
| Ток эндолимфы направлен от ампуллы | Нистагм направлен влево, горизонтальный, короткий | Ток эндолимфы направлен от ампуллы | Нистагм направлен вправо, горизонтальный, короткий |
| Правый фронтальный канал | Левый фронтальный канал | ||
| Ток эндолимфы направлен к ампулле | Нистагм направлен влево, ротаторный | Ток эндолимфы направлен к ампулле | Нистагм направлен вправо, ротаторный |
| Ток эндолимфы направлен от ампуллы | Нистагм направлен вправо, ротаторный | Ток эндолимфы направлен от ампуллы | Нистагм направлен влево, ротаторный |
Класификация нистагмов. Различают нистагмы: а) экзогенный и б) эндогенный. В основе такого разделения лежит генетический фактор. Экзогенным называется такой нистагм, который вызывается внешними причинами, например: вращением. К эндогенным нистагмам относятся те его формы, которые не имеют никакой зависимости от внешних причин, а обусловлены большей частью патологическими изменениями в самом лабиринте.
а) Экзогенный нистагм. Нистагм от вращения. Как было указано, одним из внешних раздражителей полукружных каналов является вращение (угловое ускорение) головы или всего тела. При этом возникает ток эндолимфы, отклонение купулы, вследствие чего появляется нистагм. Вращение можно назвать физиологическим раздражителем, так как повороты головы и тела происходят повседневно в нашей жизни. На обычные повороты головы или тела глаза хотя и отвечают нистагменной реакцией, но чрезвычайно слабо, и эта реакция может быть уловлена посредством нистагмографии или, менее точно, простым ощупыванием пальцами экспериментатора, а на животных при помощи методики, применявшейся Ундрицем. Другое дело, если угловое ускорение сделать необычным для человека, например: катание на каруселях, вальсирование, вращение на кресле Barany - тогда более сильный раздражитель вызовет и более отчетливое подергивание глаз. Посмотрим, что произойдет с эндолимфой при вращении на кресле Barany. Предположим, что мы вращаем человека в сидячем положении вправо. Тогда эндолимфа правого горизонтального канала в начале вращения, то - есть в тот момент, когда из состояния покоя мы перешли к вращению и, следовательно, придали исследуемому определенное угловое ускорение, по инерции будет отставать в движении от стенок перепончатого канала и будет перемещаться в сторону, противоположную вращению, т.е. от простого конца к ампулле. Согласно закону Ewald'a, это движение эндолимфы вызовет горизонтальный нистагм вправо. Одновременно в левом горизонтальном канале движение эндолимфы будет иное, а именно в первый момент вращения эндолимфа потечет от ампуллы к простому концу, что по закону Ewald'a должно вызвать нистагм тоже вправо и тоже горизонтальный. Оба импульса, исходящие из двух полукружных каналов, слагаются, в результате чего в начале вращения вправо возникает горизонтальный нистагм вправо. Таким образом, законность, выведенная для зрительного нистагма, сохраняет свою силу и для лабиринтного: нистагм своим быстрым компонентом всегда направлен в сторону движения (вращения) и происходит в плоскости движения (в данном случае горизонтальной).
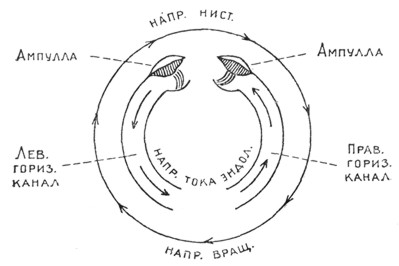 |
Рис. 4. Схема движения эндолимфы в полукружных каналах в связи с направлением нистагма. |
Описанный только что нистагм практически трудно заметить, так как исследуемый вращается. Его можно изучить лишь посредством записывающих приборов - нистагмографов.1 Если мы будем продолжать вращение исследуемого, то появившийся нистагм скоро прекратится, и глаза во время самого вращения будут находиться в состоянии покоя. Это объясняется тем, что за время вращения эндолимфа в силу своей вязкости слипается со стенками перепончатого канала и приобретает такую же скорость движения, как и стенки канала, и, следовательно, эндолимфа по каналу долго скользить не может. Как только мы остановим вращение (отрицательное ускорение), стенки канала будут находиться в состоянии покоя, эндолимфа же по инерции будет продолжать первоначальное движение вправо. Тогда в правом канале эндолимфа потечет от ампуллы к простому концу, а в левом - от простого конца к ампулле, что по закону Ewald'a вызовет горизонтальный нистагм влево (рис. 4).
Итак, после остановки вращения нистагм должен происходить в сторону, противоположную вращению. Такого рода нистагм, в отличие от нистагма, возникающего в начале вращения, называется посленистагмом. В большинстве случаев этот тип нистагма бывает более продолжителен, чем нистагм, получаемый в начале вращения. Это явление объясняется следующей гипотезой. В начале вращения, напр., вправо, происходит раздражение мозгового центра, предназначенного для нистагма, направленного в сторону вращения (Barany), но одновременно, по мнению Воячека, происходит, повидимому, и некоторое возбуждение второго центра, заведующего нистагмом в противоположную сторону, т.е. влево, и таким образом, в то время как центр нистагма вправо во время самого вращения успевает израсходовать энергию для вызывания этого нистагма, второй центр в период вращения будет находиться еще в стадии небольшого возбуждения. При остановке вращения произойдет раздражение центра нистагма влево, наслаивающееся на имеющееся уже возбуждение этого центра, и, следовательно, последний, в отличие от центра нистагма вправо, получит после остановки вращения - возбуждение плюс раздражение. Такое перераздражение центра и объясняет, почему при остановке вращения посленистагм бывает большей продолжительности, чем нистагм, вызванный первым ускорением в начале вращения. Посленистагм считается более удобным для наблюдения, т. к. исследуемый находится в состоянии покоя, и практически клиницисты судят о функции полукружных каналов большею частью по посленистагму. Необходимо помнить, что в посленистагме после вращения вправо главное участие принимает левый лабиринт, после вращения влево, наоборот - правый. Это положение объясняется тем же законом Ewald'a.
Известно, что ток эндолимфы в горизонтальном канале к ампулле вызывает нистагм, более продолжительный и качественно наиболее резкий, чем ток от ампуллы. При остановке вращения вправо в левом канале получается ток к ампулле, а в правом - от ампуллы и, следовательно, продолжительность нистагма и его четкость зависят преимущественно от левого полукружного канала. После вращения влево картина будет обратная. Обычно при нормальных лабиринтах постнистагмические подергивания глаз после остановки вращения вправо и влево бывают одинаковой продолжительности, а когда один лабиринт выключен (парализован), продолжительность посленистагма будет неодинакова при правом и левом вращении. Возьмем для примера случай свежего паралича правого лабиринта и посмотрим, какой результат должен получиться после вращения в обе стороны. Вполне понятно, что при рассуждениях о токах эндолимфы мы должны иметь в виду один только левый полукружный канал. После вращения вправо в левом канале эндолимфа потечет к ампулле, что по закону Ewald'a вызовет продолжительный и отчетливый нистагм. После же вращения влево в левом канале произойдет ток эндолимфы от ампуллы и это вызовет нистагм короткой продолжительности и слабо выраженный. Таким образом, контраст продолжительности и четкости нистагма после правого и левого вращения говорит о параличе полукружных каналов одной стороны. Противоположная картина наблюдается в случае раздражения одного из лабиринтов. Тогда нистагм, вызванный вращением и зависящий от раздражения больного лабиринта, будет более отчетливым и продолжительным, чем нистагм, полученный от лабиринта здоровой стороны.
Средняя нормальная продолжительность горизонтального нистагма равна 25-35". При горизонтальном вращении эндолимфа фронтального и сагиттального каналов движется слабо, поэтому эти каналы раздражаются тогда в весьма умеренной степени. Если же исследуемый наклонит голову вперед на 90°, то в плоскость вращения установятся преимущественно фронтальные2 каналы, и тогда - после остановки вращения - появляется постнистагм по закону Ewald'a, ротаторного типа и направленный в сторону, противоположную вращению. Если производить вращение исследуемого с наклоненной на 90° к плечу головой, то в плоскости вращения будут установлены преимущественно сагиттальные каналы, и тогда после остановки вращения появится вертикальный постнистагм, направленный в сторону, противоположную вращению. Продолжительность постнистагмов, полученных с фронтальных и сагиттальных каналов, бывает короче постнистагма, полученного с горизонтальных каналов, и в среднем она равна 10-15 секундам.
Бывают нистагмы, имеющие в своей основе раздражитель искусственный, не физиологический. Оказывается, что ток эндолимфы полукружных каналов можно получить не только специфическим раздражителем, угловым ускорением, но и рядом искусственных мер, например: охлаждением, прогреванием каналов, пропусканием гальванического тока, повышением и понижением давления в канале. По характеру применения того или иного искусственного раздражителя и различают нистагмы: калорический, механический, прессорный и гальванический.
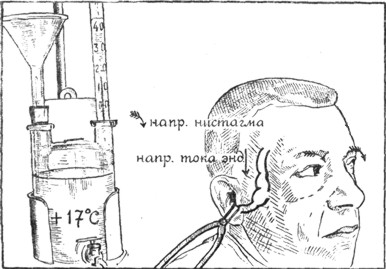 |
Рис. 5. Схема калорического нистагма: вливание прибором Brunings'a холодной воды (+17°С). Простая стрелка показывает направление тока эндолимфы в правом вертикальном канале, что вызывает ротаторный нистагм влево (перистые стрелки обозначают направление нистагма). В горизонтальном канале движения эндолимфы нет или оно незначительное (по Воячеку). |
Калорический нистагм. Хотя возможность раздражения лабиринта от вливания в ухо воды была известна давно, но только с 1904 года, после работы Barany, калорический нистагм был введен в систему вестибулометрии. Если человеку влить, например, в правое ухо холодную воду, то произойдет охлаждение наружного слухового прохода, барабанной перепонки, барабанной полости и, наконец, боковой стенки лабиринта. Вследствие охлаждения некоторого участка полукружного канала произойдет ток эндолимфы, зависящий от опускания вниз охлажденных частиц эндолимфы. При вертикальном (нормальном) положении исследуемого высшую точку лабиринта образует дуга фронтального канала и, следовательно, при калоризации ток эндолимфы в правом канале пойдет по длине канала от простого конца к ампулле, что по закону Ewald'a вызовет ротаторный нистагм влево (рис. 5). Одновременно в правом горизонтальном канале тоже произойдет опускание эндолимфы, но этот ток будет идти поперек или почти поперек оси канала, от верхней стенки к нижней. Такое движение эндолимфы не по оси канала не может вызвать значительного отклонения купулы, следовательно, результирующий нистагм будет преимущественно ротаторным, т.е. во фронтальной плоскости. Другое дело, если мы после вливания наклоним голову исследуемого на левое плечо на 90°,- тогда высшей точкой лабиринта будет дуга правого горизонтального канала, и эндолимфа потечет по оси этого канала от простого конца к ампулле, что вызовет появление горизонтального нистагма вправо (закон Ewald'a). Если же после охлаждения наклонить голову на правое плечо, то наивысшей точкой будет ампулла горизонтального канала, и эндолимфа тогда потечет от ампуллы к простому концу, что вызовет горизонтальный нистагм влево (рис. 6). При вливании в ухо воды, температура которой выше температуры тела, будут действовать обратные физические законы, и тогда направление тока эндолимфы, а следовательно, направление нистагма становится обратным; например, при вливании такой воды в правое ухо человека, сидящего в нормальном вертикальном положении, появится ротаторный нистагм вправо и т.д.
Обычно калоризация уха для вызывания нистагма производится холодной водой температуры между 4-15° и + 30°С и теплой водой температуры +430C.3 Чаще пользуются холодной водой, теплая же калоризация применяется только в тех случаях, когда у больного имеется так наз. самопроизвольный (спонтанный эндогенный) нистагм в здоровую сторону. Тогда при горячей калоризации может произойти изменение самопроизвольного нистагма, т.е. он может совсем прекратиться и, больше того, может появиться калорический нистагм в больную сторону. Понятно, что это произойдет только в том случае, если исследуемый больной лабиринт еще функционирует. Если лабиринт полностью парализован, вливание горячей воды не даст нистагма и не окажет никакого влияния на имеющийся самопроизвольный эндогенный нистагм.
Как разновидность калорической пробы в некоторых клиниках применяют иногда способ минимальных раздражений, заключающийся в том, что в ухо вливают небольшие порции воды. Этот способ рекомендуется в тех случаях, когда у исследуемого предполагается здоровый или раздраженный лабиринт и, следовательно, для исследования функции нет надобности в применении сильного раздражителя (такого взгляда придерживается Kobrak).
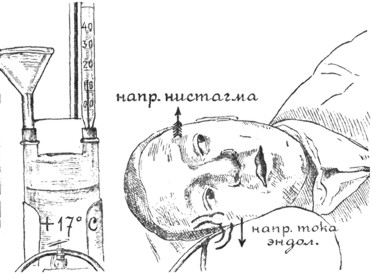 |
Рис. 6. То же, что и на рис. 5; при наклонении головы на 90° к правому плечу эндолимфа правого лабиринта движется только в горизонтальном канале, от ампуллы к гладкому концу, отчего получается (слабый) горизонтальный нистагм налево (по Воячеку). |
Прессорный нистагм. Этот вид нистагма у большинства людей получить не удается, так как он возможен только при врожденной дегисценции или узуре костной стенки полукружного канала при хронической оторрее. Из анатомии известно, что дуга горизонтального канала выстоит в antrum сосцевидного отростка. При хроническом гнойном воспалении среднего уха кариозный процесс может образовать свищ в этом месте костного горизонтального канала, и тогда при сгущении воздуха в наружном слуховом проходе давление передается на перепончатый канал и перемещает эндолимфу от простого конца к ампулле, что, по закону Ewald'a, вызывает горизонтальный нистагм в сторону исследуемого уха. При разрежении давления в наружном слуховом проходе ток эндолимфы пойдет от ампуллы к простому концу, в результате чего появится горизонтальный нистагм в противоположную сторону.
Гальванический нистагм (Hitzig, 1870 г.). Этот вид нистагма особого практического значения не имеет, т. к. существует предположение, что при пропускании гальванического тока получаемый нистагм не зависит от появления токов эндолимфы, а обусловлен раздражением самого ствола вестибулярного нерва.
б) Эндогенный, спонтанный, самопроизвольный нистагм. Если у человека наблюдается нистагм без применения приведенных выше искусственных и физиологических раздражителей, а обусловленный причинами эндогенного характера, то такой нистагм называется спонтанным или самопроизвольным. Лабиринтный спонтанный нистагм бывает в тех случаях, когда полукружные каналы на одной стороне раздражены или угнетены сильнее, чем на другой. В случае раздражения, напр., правого лабиринта нистагм бывает в правую сторону и, наоборот, в случае угнетения того же лабиринта появляется спонтанный нистагм в левую, здоровую сторону. Наиболее простое объяснение сущности спонтанного нистагма заключается и следующем гипотетическом предположении. Нормальные лабиринты посылают равные по силе тонические импульсы к мышцам глаз, и каждый из лабиринтов стремится вызвать нистагм в свою сторону, но так как правый и левый лабиринты взаимно - антагонистичны, то в норме глаза находятся в состоянии тонического равновесия, и никакого нистагма не бывает. Представим себе, что правое ухо заболевает гнойным отитом, и может случиться, что токсины, а может быть и гной, проникнут во внутреннее ухо. Тогда в первые часы (дни) произойдет раздражение токсинами или гноем ампуллярных окончаний правого вестибулярного нерва, которые значительно сильнее воспримут раздражение от постоянно существующего слабого физиологического тока эндолимфы (Breuer), чем не раздраженные нервные окончания здорового левого лабиринта. Следствием этого будет функциональный перевес правого лабиринта над левым, в результате чего наступит спонтанный нистагм в правую, больную сторону. Обычно такой нистагм в стадии раздражения бывает непродолжительное время, так как эту стадию скоро сменяет стадия паралича, и тогда картина спонтанного нистагма изменяется. При параличе правого лабиринта здоровый левый лабиринт, потеряв своего антагониста, начинает проявлять себя свободно, и именно физиологический ток эндолимфы вызывает спонтанный нистагм в свою левую, здоровую сторону. Такого рода нистагм может существовать более продолжительное время (месяцами), чем спонтанный нистагм в стадии раздражения. Он продолжается до тех пор, пока не наступит компенсация других органов, участвующих в мышечной координации (центр, нервной системы, зрения, мышечного, суставного чувства и т.д.).
При наблюдении спонтанного нистагма очень легко впасть в ошибку, т. к. встречаются особые формы спонтанного нистагма, не связанные с заболеванием полукружных каналов, но могущие "симулировать" лабиринтный спонтанный нистагм. К таким нистагмам относятся: физиологический нистагм, интенционный нистагм, зависящий от неправильной аккомодации зрительного аппарата и, наконец, нистагм, обусловленный заболеванием центральной нервной системы.
Физиологический спонтанный нистагм отличается от патологического лабиринтного тем, что он, хотя и ритмический, т.е. состоит из быстрого и медленного компонентов, но наблюдается всегда только при крайнем отведении глаз, чрезвычайно слабо выражен и бывает одинаковой силы при взгляде в ту и другую стороны.
Интенционный или установочный нистагм бывает тоже ритмичным, но очень непродолжительным, всего 2-3 секунды, и зависит, невидимому, от временной контрактуры глазных мышц, возникающей при попытке крайнего отведения глаз.
Нистагм, зависящий от аномалии зрительного аппарата, отличается от лабиринтного спонтанного нистагма тем, что может быть не ритмичным, т.е. не имеет быстрого компонента - глаза ундулируют, движения их напоминают качание маятника, т.е. движение в ту и другую стороны происходит с одинаковой скоростью.
Нистагм, обусловленный заболеванием центральной нервной системы (напр., мозжечка), тоже довольно заметно отличается от лабиринтного. Такой нистагм обычно бывает ритмичным, но ритм его неравномерен, беспорядочен, т.е. быстрый компонент может неожиданно заменяться медленным и наоборот. Если перевести на музыкальный ритм, то лабиринтный нистагм бьет по счету 3/4, а "нервный" нистагм по счету 3/4, 1/2, 3/4, 1/1, 1/2 и т.д.
Если дифференциация спонтанного нистагма все - таки окажется трудной, то генез его можно проверить при помощи нистагмографов или посредством вышеизложенных методов исследования лабиринтной функции. При отсутствии каких - либо уклонений от нормы со стороны лабиринта спонтанный нистагм приходится отнести к нелабиринтным.
Заканчивая главу о нистагме, необходимо сделать критическую оценку всех перечисленных способов исследования.
Исследование спонтанного нистагма считается наиболее нежным и чувствительным. Клинически это исследование имеет большое значение, т. к. до некоторой степени позволяет судить о стороне и стадии заболевания полукружных каналов. Следующей по своей чувствительности является калорическая проба. Посредством калорической реакции можно узнать о состоянии возбудимости окончаний вестибулярного нерва, применив самый нежный искусственный раздражитель- молекулярное смещение столбика эндолимфы. Вполне понятно, что, если при этом способе получается нистагм, то говорить о сильном угнетении функции лабиринта уже не приходится; кроме того, основная ценность калорической пробы заключается еще и в том, что этот способ позволяет изолированно исследовать каждое ухо. Менее ценным является исследование полукружных каналов вращением, т. к. при этом способе одновременно раздражаются оба лабиринта. В отношении- силы раздражения вращение считается более грубым способом, чем калорическая проба. Все же и вращение на практике крайне необходимо, т. к. может случиться, что калорическая реакция будет отрицательна, вращением же удастся вызвать нистагм. Это будет говорить о том, что эндолимфа изменила свои физические свойства (напр., стала более вязкой), и нежная калорическая проба не в состоянии была вызвать смещение эндолимфы, грубый же раздражитель- вращение - имел еще возможность сместить измененную эндолимфу. Тогда вращение будет указывать, что полного паралича окончаний вестибулярного нерва еще не наступило. Еще более грубым считается исследование прессорного нистагма. Бывают случаи, когда спонтанный нистагм направлен в здоровую сторону, калоризация и вращение не дают нистагма, и, казалось бы, можно уже говорить о параличе лабиринта, прессорный же опыт дает отчетливый нистагм. В таком случае диагностировать паралич вестибулярных окончаний преждевременно. Если у такого больного исчезнет и прессорный нистагм, тогда только можно с уверенностью сказать о выключении лабиринта. Имея в основе эти положения о силе раздражителей и учитывая их практическую значимость, рекомендуют исследование полукружных каналов производить по следующей схеме (последовательности):
| Правое ухо | Левое ухо |
|
|
б) Рефлексы на мышцы конечностей и туловища. Известно, что нормальные лабиринты посылают одинаковые по силе тонические импульсы к мышцам глаз, причем правый лабиринт по своей рефлекторной дуге тонизирует те мышцы, которые заведуют медленным компонентом глаз в левую сторону (перекрестно), субкортикальные же центры дают быстрый компонент в правую сторону. Левый лабиринт действует в противоположном смысле. В норме оба лабиринта, действуя одновременно как антагонисты, удерживают глазные мышцы в состоянии тонического равновесия, и никакого нистагма не наступает. То же самое распределение лабиринтного тонуса существует и в отношении остальной мускулатуры нашего тела, а именно правый лабиринт заведует тонусом мышц, обусловливающих движение нашего тела влево, левый же лабиринт - наоборот. Если искусственно вызвать раздражение, например, правого лабиринта, то появится гипертонус мышц, заведующих движением тела влево; если же разрушить правый лабиринт, то левый, потеряв своего антагониста, даст гипертонус мышц, заведующих движением тела вправо. Таким образом, видна полная аналогия лабиринтных тонических рефлексов мышц глаз и остальных мышц тела. Особенно отчетливо эту аналогию удается наблюдать при экспериментальном исследовании функции полукружных каналов мелких животных, например, мышей. После вращения на центрофуге животное совершает ритмическое подергивание всем телом. Такое явление называется нистагмом всего тела, и быстрый компонент его, подобно нистагму глаз, после остановки всегда направлен в сторону, противоположную вращению. У птиц после вращения появляется нистагм головы. У крупных животных (кролик, кошка, собака) и у человека после вращения, как правило, ни головного нистагма, ни нистагма всего тела не бывает. Можно сказать, что у человека ритмические рефлекторные движения после раздражения лабиринтов сохранились только в отношении глазных мышц, все же остальные мышцы тела филогенетически потеряли быстрый компонент, а медленный остался в полной силе, хотя индивидуально и вариирует в больших пределах. Из сказанного понятно, что двигательный эффект этих рефлексов бывает направлен в сторону медленного компонента глазного нистагма. Какие бы ни были раздражители полукружных каналов (калоризация, вращение, компрессия, декомпрессия и т.д.), законность эта всегда сохраняет свою силу. Возьмем для примера экзогенный раздражитель - вращение: у субъекта получен нистагм вправо после вращения влево и, значит, медленный компонент будет направлен влево; тогда наиболее гипертонизированными будут те мышцы конечностей и туловища, которые производят движение влево.
Изменение тонуса мышц конечностей и туловища может быть замечено посредством специальных измерительных приборов-тонометров (Voss) или, еще проще, исследованием активных движений. Если исследуемого с правосторонним нистагмом после вращения попросить закрыть глаза и прикоснуться рукой или ногой к какому -либо неподвижному предмету, то он будет промахиваться, т.е. конечности его будут отклоняться влево от предмета. То же самое происходит и с мышцами туловища. У такого субъекта мышцы туловища левой стороны будут находиться в состоянии повышенного тонуса, и это должно вызывать нагибание туловища влево. При покойном положении исследуемый компенсирует это нагибание посредством проприоцептивных органов (глаза, мышечное, суставное, тактильное чувство и др.), но если его заставить пройти с закрытыми глазами по прямой линии, то окажется, что походка его будет неуверенная и, кроме того, он будет отходить влево от этой линии. Такое нарушение походки и отклонение от заданного направления объясняется двумя причинами: 1) повышенным тонусом мышц нижних конечностей, заведующих движением влево (абдукторы левой ноги и аддукторы правой), и 2) повышенным тонусом левосторонней мускулатуры туловища, обусловливающим смещение центра тяжести влево ; чтобы сохранить равновесие, исследуемый, отклоняясь влево от прямой линии, стремится тем самым установить центр тяжести в нормальное положение.
Возникает вопрос, какова же функциональная ценность описанных реакций отклонения. Одно из возможных объяснений состоит в следующем. Будем вращать исследуемого в кресле Barany влево, а затем остановим вращение. Тогда исследуемый будет чувствовать противовращение. Ему будет казаться, что он сам вращается вправо, а все окружающие предметы движутся влево, и что он должен упасть, если не будет следовать за убегающими от него предметами. Чтобы защитить себя от падения, он будет стремиться как бы догнать удаляющиеся влево предметы ; следовательно, его туловище и конечности будут отклоняться влево. На этом основании эти рефлексы называются защитными (от падения). Защитные рефлексы появляются независимо от того, будет ли исследуемый видеть объективные окружающие предметы в кажущемся движении; они возникают и в том случае, если исследуемый закроет глаза, но тогда в его представлении будут двигаться так называемые субъективные, мыслимые им предметы (субъективное головокружение).
На практике исследование тонических лабиринтных рефлексов на мышцы производится с помощью так наз. указательной пробы Вагапу и реакции падения.
в) Рефлексы на гладкую мускулатуру (вегетативные) При раздражении полукружных каналов появляется ряд рефлексов со стороны вегетативной нервной системы. Сюда относятся: холодный пот, гусиная кожа, изменение окраски кожных покровов (побледнение или покраснение), тошнота, рвота и дефекация. Эти рефлексы проходят по своей рефлекторной дуге, имеющей афферентным нейроном n. vestibularis, коннекторным - повидимому, corpus striatum и эфферентным- периферические нервы вегетативной системы. Вегетативные рефлексы имеют большое практическое значение, т. к. их легко можно наблюдать при обычных способах исследования. Из всех рефлексов наиболее частыми и наиболее мучительными являются тошнота и рвота (триада лабиринтных поражений - глухота, головокружение и тошнота).
Помимо упомянутых рефлексов, существует еще группа вегетативных реакций, имеющая пока только теоретическое значение. Так, Spiegel, Demetriades, Перекалин, Schon, Goldberg, Wotzilka, Stein, Benesi, Szasz и др. наблюдали при раздражении полукружных каналов изменение кровяного давления, сердечной и дыхательной функций, перистальтики кишек и зрачковой реакции. Из этой группы реакций наиболее изученной является сосудистая. По данным Перекалина, раздражение полукружных каналов вызывает начальное падение и последующий подъем кровяного давления. После адренализации (симпатикотропный яд) животного эта реакция исчезает. Это говорит о том, что лабиринтный вегетативный рефлекс на сосудистую систему идет по симпатическим путям. Какой из симпатических нервов является эфферентным нейроном рефлекторной дуги, остается вопросом, недостаточно еще выясненным. Но, все - таки в отношении, например, кровяного давления, можно предполагать, что таковым является, повидимому, n. splanchnicus, т. к. перерезывание n. sympathici cervicalis и n. vagi заметного влияния на сосудистый рефлекс не оказывает.
Сосудистый рефлекс не лишен и некоторого практического значения. Лапин и Зброжек, исследуя кровяное давление у людей (по Riva-Rocci) при раздражении лабиринта, получали изменение кровяного давления аналогично экспериментальным данным Перекалина. На основании имеющегося экспериментального материала можно сказать, что пока мы имеем только факты, с достоверностью говорящие о влиянии полукружных каналов на функции вегетативной нервной системы. Какой из компонентов вегетативной нервной системы (парасимпатическая или симпатическая нервная система) играют здесь преимущественную роль, сказать трудно. Клинические наблюдения показывают, что законности в этом отношении, повидимому, не существует, и характер лабиринтной вегетативной реакции зависит от вегетативной конструкции человека, и именно у ваготоников может быть своя реакция, у симпатикотоников же - обратная. После вращения, например, мы видим, что одни исследуемые очень бурно реагируют в смысле вегетативных реакций, другие - наоборот, у одних наступает покраснение лица, у других - побледнение. Только упрощение и совершенствование методики исследования вегетативных рефлексов позволит в дальнейшем ввести в практику регистрацию лабиринтных вегетативных рефлексов.
|
The requested URL /down.htm was not found on this server.